原油備蓄設備からの金属腐食菌のスクリーニング
原油備蓄タンク等の試料から多種多様な嫌気性微生物を幅広く分離・培養し、それら微生物の鉄腐食能の有無を検討することで、鉄腐食菌の分離を目指しました。
その一環で、同様の試料から、偏性嫌気性水素資化性微生物を幅広く分離し、それらの微生物の鉄腐食能を調べました。これは、昔からの命題「嫌気的環境において鉄から化学的に発生する水素を食べる微生物が腐食を引き起こす」の検証を目的としています。
さらに、鉄を唯一の電子供与体とした嫌気的な集積培養を行い、鉄腐食菌の直接的な分離を目指しました。
a.嫌気性菌の網羅的な分離と鉄腐食能の検討
原油備蓄施設、原油生産施設、天然ガス製造所等から採取した試料(【図1】)から嫌気微生物を分離しました。
【図1】微生物の分離・培養を目的として収集した資料

原油 (3 試料)
MIC1、MIC3、MIC5

ドレイン水 (6 試料)→ドレイン水
MIC2、MIC8、MIC10、MIC12、MIC13、MIC14

鉄サビ (2 試料)
MIC4、MIC6

堆積物 (3 試料)
MIC7、MIC9、MIC11
MIC1、2、8:秋田県 │ MIC3、4:宮城県 │ MIC5、6:秋田県
MIC7、9、11、13:千葉県 │ MIC10:新潟県 │ MIC12:大阪府
嫌気性微生物は、しばしばコロニー形成が困難なため、主に液体培地を用いて集積培養を行いました。培地は水素資化性及び有機酸資化性の硫酸塩還元菌及びメタン生成古細菌を対象としたものを使用し、培養温度は25℃としました。嫌気培養瓶内で寒天によるコロニー化(混釈法)を行い、コロニーの形成が認められたものについては、液体培地に移しました。コロニー生成が認められないものについては、限界希釈法による分離を行いました。分離した微生物については、16S rRNA遺伝子配列の決定による簡易同定を行いました。
分離培養の結果、64属78種の微生物株を獲得できました。微生物株が得られた分類群を系統樹(【図2】)に示しました。嫌気的に増殖を行うことが可能な中温性微生物に関する系統の大部分を得ることができました。分離源である原油スラッジや配管等には様々な種類の微生物が存在し、潜在的に様々な微生物が増殖する可能性をもっている環境であるといえます。16S rRNA遺伝子配列の相同性として98 %以下を別種であるとした場合に、既知種と一致しないものが119株に上り、新規な微生物種が多く分離できたことが判明しました。分離した全微生物の16S rRNA遺伝子配列に基づいて、できる限り多種多様な30株(【図2】の赤色字)を選抜して鉄腐食能を検討しました。
【図2】分離した微生物の系統と鉄腐食能検討株

分離した微生物の鉄腐食能を検討するために、鉄顆粒による培養を行いました。培地は、人工海水培地に2.7Mの鉄顆粒及び硫酸塩や硝酸塩を添加したものを使用しました。前培養は分離に使用した培地を使用し、すべて25℃で培養し、各条件下で鉄腐食が起こるか否かを検討しました。メタン生成古細菌に関しては硫酸塩添加区のみ行いました。非生物的な腐食として未植菌区を陰性対照に設定しました。検討した30株のうち、代表的なものを【図3】に示しました。未植菌区では、硫酸塩又は硝酸塩存在下、それぞれ平均で4.6mMと2.8mMの鉄が化学的に腐食されることがわかりました。
硫酸塩が存在する場合、微生物株を添加したもので明らかな鉄腐食を誘導したのは、Methanococcus sp. MIC11-43 のみでした。硫酸塩還元菌であるDesulfotomaculum sp. MIC4-22、Desulfobacter curvatus MIC4-9、Desulfovibrio dechloracetivorans MIC5-15では、非生物的な鉄腐食以上の鉄腐食は起こりませんでした。一方、硝酸塩が存在する場合、Desulfotomaculum sp. MIC4-22、Prolixybacter sp. MIC1-1、Desulfobacter curvatus MIC4-9、Malonomonas sp. MIC11-27、Thiomicrospira sp. MIC5-2、Shewanella sp. MIC5-4及びThiobacillus sp. MIC3p-35について嫌気環境下における鉄腐食が確認できました。Proteobacteriaに属するものが多く腐食能をもっている傾向を示しましたが、腐食能をもつ微生物に特筆すべき系統学的な特徴は見受けられませんでした。
| 分離同定した微生物 | 硫酸塩存在下における 鉄腐食量 |
硝酸塩存在下における 鉄腐食量 |
|---|---|---|
| Archaea (古細菌) | ||
| Euryarchaeota | ||
| Methanococcus sp. MIC11-43 | 8.6 mM (灰色) | ─ |
| Methanolobus sp. MIC11-52 | 5.4 mM (変化なし) | ─ |
| Bacteria (細菌) | ||
| Firmicutes | ||
| Clostridium sp. MIC7-2 | 3.5 mM (変化なし) | 3.2 mM (変化なし) |
| Fusibacter sp. MIC7-3 | 1.4 mM (変化なし) | 2.4 mM (変化なし) |
| Acetobacetrium sp. MIC1-12 | 2.2 mM (変化なし) | 2.3 mM (変化なし) |
| Proteobacteria | ||
| Desulfovibrio sp. MIC5-15 | 4.8 mM (変化なし) | ─ |
| Malonomonas sp. MIC11-27 | 2.5 mM (変化なし) | 9.0 mM (茶褐色) |
| Thiomicrospira sp. MIC5-2 | 5.1 mM (変化なし) | 8.7 mM (茶褐色) |
| Shewanalla sp. MIC4-4 | 5.2 mM (変化なし) | 20.1 mM (黒色) |
| その他 | ||
| Geotoga sp. MIC4-5 | 3.2 mM (変化なし) | 5.6 mM |
| Dethiosulfovibrio sp. MIC9-28 | ─ | ─ |
| Marinilabilia sp. MIC5-40 | 2.4 mM (変化なし) | 2.1 mM (変化なし) |
| Bacteroide -like bacterium MIC1-1 | 1.7 mM (変化なし) | 10.4 mM (黒色) |
| Control (未植菌) | ||
| Not inoculation | 4.6 mM (変化なし) | 2.8 mM (変化なし) |
* 数値は溶出鉄濃度、かっこ内は培養後の鉄顆粒の状態を示す。
以上をまとめると、以下のようになります。
- 試料から多種多様な微生物を分離・培養し、それらの金属腐食能の有無を検討することで、微生物腐食の原因菌の系統学的な分布を調べました。
- Euryarchaeota、Proteobacteria、Firmicutes、Bacteroidetes等に属する多様な微生物を分離し、64属78種に系統学的に分類できました。
- Methanococcales、Delta-proteobacteria、Epsilon-proteobacteria、Gamma-proteobacteria、Bacteroidetes、Desulfotomaculumに分類される微生物が鉄腐食能を示しました。それぞれの鉄腐食は、現象的に様々でした。
文献 1:Iron-corroding methanogen isolated from a crude-oil storage tank. Appl Environ Microbiol. 2010 Mar;76(6):1783-8.
文献 2:Iron corrosion induced by nonhydrogenotrophic nitrate-reducing Prolixibacter sp. strain MIC1-1. Appl Environ Microbiol. 2015 Mar;81(5):1839-46.
文献 3:鉄を電子供与体として利用可能な嫌気性微生物の集積培養物による鉄腐食. 材料と環境 2010 59, 298-304.
b.偏性嫌気性水素資化性菌の分離と鉄腐食能の検討
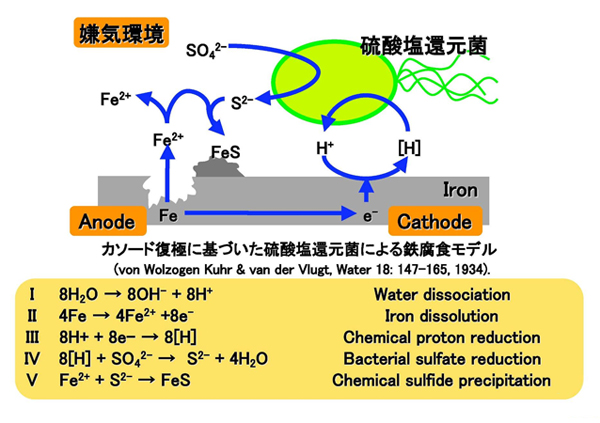
鉄が嫌気条件下の水に浸かった時に起こる化学的な鉄の酸化(Fe → Fe2+ + 2e-)は、水由来のプロトン(H+)によって促進され、分子状水素が発生する(Fe + 2H2O → Fe2+ + H2 + 2OH-)。この化学的に発生した水素は、カソード水素(cathodic hydrogen)と呼ばれています。Von Wolzogen KuehrとVan der Vlugtは、1934年に「鉄が水に接したときに形成される"カソード水素"や"水素膜(hydrogen film)"を微生物が消費した結果として微生物腐食が起こる」という仮説を立てました。これは、"カソード復極説(Cathodic depolarization Theory)"と呼ばれ、微生物による鉄腐食機構として広く知られています(【図4】)。
しかし、これまでに本仮説を直接的に証明する実験結果は得られておらず、疑問が持たれ続けてきたことも事実です。
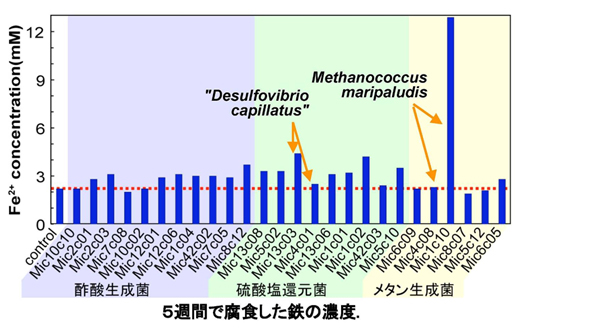
嫌気環境でのカソード復極による微生物腐食を検証することを目的として、偏性嫌気性水素資化性菌を分離し、金属腐食能の検討を行いました。Firmicutesに属する酢酸生成細菌11株、Delta-proteobacteriaに属する硫酸塩還元細菌9株及びArchaeaに属するメタン生成古細菌6株を使用して、鉄腐食を検討しました。鉄腐食試験として、鉄顆粒存在下で25℃にて5週間培養した後の培養液中の鉄イオン濃度を測定しました(【図5】)。非生物的な腐食として未植菌区も検討し、5週間で平均2.2mMの化学的腐食が起こりました(【図5】の赤点線)。
非生物的な腐食と比較して強い腐食能を示したのは、Methanococcus maripaludis Mic1c10のみでした。一方、16S rRNA遺伝子配列的に完全に同種であると判断できるM. maripaludis Mic4c08は、全く腐食能を示しませんでした。この結果は、同じM. maripaludisであっても、腐食能に関して株レベルで差異があることを示しています。M. maripaludis Mic1c10以外では、一部の硫酸塩還元菌(Desulfovibrio capillatus Mic13c03とDesulfovibrio dechloracetivorans Mic1c02)が非生物的な腐食と比較して約2倍程度の鉄腐食能を示したのみでした。これらの培養液中では水素が枯渇した状態であり、化学的に発生した水素はそれぞれの微生物によって常に消費されていたことを示しています。一方M. maripaludis Mic1c10では化学的に発生する水素の数倍量の水素が発生していました。
以上の結果、カソード復極による微生物腐食と考えられる現象が硫酸塩還元菌等で確認はできましたが、当該微生物腐食は、非生物的(化学的)な腐食と比べて最大で2倍程度であり、嫌気環境下において深刻な微生物腐食とはならないことが示唆されました。一方、偏性嫌気性水素資化性菌であるM. maripaludis Mic1c10で起こった鉄腐食は、これらカソード復極に基づいた微生物腐食と比べて非常に激しく、能動的なM. maripaludis Mic1c10の関与が考えられました。また、通常では考えられないような水素発生が示されたことから、本腐食はカソード復極による腐食とは異なる全く新しい微生物腐食機構である可能性を示唆しました。
文献 4:Iron corrosion activity of anaerobic hydrogen-consuming microorganisms isolated from oil facilities. J Biosci Bioeng. 2010 Oct;110(4):426-30.
c.鉄腐食菌の直接濃縮
嫌気条件下で鉄を腐食させる微生物を濃縮するため、金属鉄を唯一の電子供与体として嫌気性菌を集積濃縮しました。全国各地の原油タンク底水を、鉄顆粒を唯一の電子供与体とする培地(Fe培地)に接種し、N2/CO2ガス下で37℃で1週間嫌気培養しました。1週間後、培養液を新しい培地に植え継ぎ、この操作を繰り返しました。最終的に微生物を分離するため、寒天を用いて固形化した培地上で水素を唯一の電子供与体として培養し、コロニー形成を行いました。以上の結果、鹿児島県及び大阪府由来のサンプルから、それぞれ、KA1株(NBRC102054)及びOS7株を得ることが出来ました。これらの株の16S rRNA遺伝子配列は、Methanococcus maripaludis JJTのそれと100 %一致し、両株ともM. maripaludis であると同定しました(【図6】)。

近年、H. T. Dinhらは、微生物腐食に大きく関与する微生物として、海洋堆積物から硫酸塩還元菌Desulfobacterium sp. IS4株、Desulfomicrobium sp. IS5株、Desulfovibrio sp. HS2株及びメタン生成古細菌Methanobacterium sp. IM1株の4株を分離しました(H. T. et al. Nature 427: 829-832. 2004)。また、硫酸塩還元菌は、その生育に伴った硫化水素の発生により金属を著しく腐食させるため、微生物腐食の代表的な原因菌として扱われ、研究対象とされてきました。メタン生成古細菌についても、嫌気的有機物分解において最終産物の除去を担う微生物群として重要であることから、微生物腐食にも強く関与している可能性が示唆されてきました。今回、原油備蓄タンク底水から分離した鉄腐食菌もM. maripaludisであったこと、また、他の方法で分離した鉄腐食性メタン生成古細菌がいずれもM.maripaludisと分類された(前記b.項)ことから、この仲間が原油タンク内部の腐食に関わっている可能性が示されました。
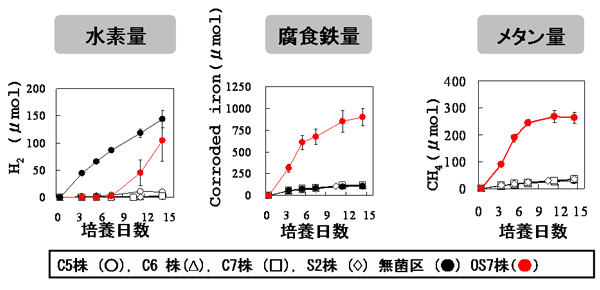
鉄腐食能を調べるために、分離株をFe培地で培養し、培養期間中の水素量、メタン量、腐食鉄量を測定しました。無植菌区は、嫌気的無菌的条件での腐食を調べるために使用しました。嫌気的無菌的条件でも、化学的な鉄の酸化が起こり、わずかな量の水素が生成し、メタンの発生は認められませんでした。一方、OS7株の培養液からは、培養初期には水素発生が認められず、メタン生成が認められました。メタン生成量は、無菌的に発生する化学的酸化による水素生成量から予想されるメタン生成量(メタン生成古細菌は、4 molのH2から1 molのCH4を生成する:4H2 + CO2 → CH4 + H2O)よりもはるかに大きい値でした。メタン生成量が増加している間は水素が検出されませんが、メタン生成の増加が起こらなくなってからは水素が検出され始めました。OS7株による鉄腐食は、培養14日目で無菌区のそれと比較して約8倍高いことが分かりました。KA1株でも同様の結果が得られました。 様々なカルチャー・コレクションに保存されている4株のM. maripaludis(C5株、C6 株、C7株、S2株)についても同様に鉄腐食能を検討しました。これらの株でのメタン生成量は、無菌的に発生する化学的酸化による水素生成量から予想されるメタン生成量と一致しました。このことから、OS7株及びKA1株以外のM.maripaludisは、鉄腐食を誘導しないと結論しました(【図7】)。
文献 1:Iron-corroding methanogen isolated from a crude-oil storage tank. Appl Environ Microbiol. 2010 Mar;76(6):1783-8.
d.鉄腐食性メタン生成古細菌と硫酸塩還元菌とによる協調的な鉄腐食促進作用
自然界ではある微生物が単独で微生物腐食を起こしているとは限りません。それを示唆する一例を発見しました。鉄片(1 cm平方、0.1 cm厚)をN2/CO2(4/1)下で鉄腐食性メタン生成古細菌M. maripaludis OS7株単独で培養した場合に比べて、鉄腐食能を持たない硫酸塩還元菌を共培養した場合により激しい金属腐食が認められました(【図8】)。

この鉄片を切断し、試料断面を走査型電子顕微鏡を用いて観察しました。鉄腐食能をもたない硫酸塩還元菌とM. maripaludis OS7株の共培養で、鉄片の肉厚が著しく減少していることが分かります(【図9】)。

腐食断面についてエネルギー分散型X線装置付き走査型電子顕微鏡を使用して、M. maripaludis存在下での腐食、及び鉄腐食能をもたない硫酸塩還元菌とM. maripaludis OS7株を共培養したときの腐食について、腐食断面の元素分布を調べました。M. maripaludis存在下での腐食では、C、O及びFeに富んでいました(【図10】)。
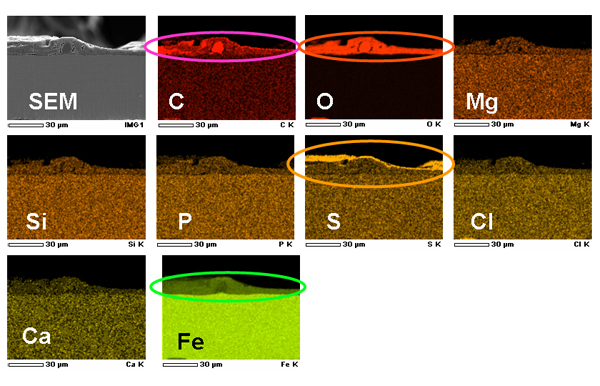
一方、鉄腐食能をもたない硫酸塩還元菌とM. maripaludis OS7株の共培養での腐食では、それに加え、Sが強く検出されました(【図11】)。腐食断面をX線解析した結果、両域ともFeCO3に富むことが分かりました。

お問い合わせ
- 独立行政法人製品評価技術基盤機構 バイオテクノロジーセンター バイオ技術評価・開発課(かずさ)
-
TEL:0438-20-5764
住所:〒292-0818 千葉県木更津市かずさ鎌足2-5-8 地図
お問い合わせフォームへ